Pohlia
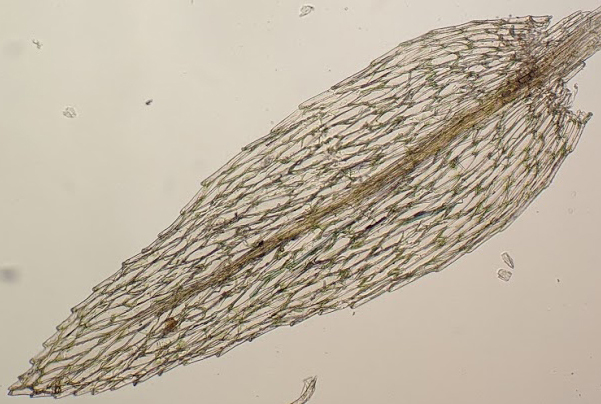
Dioicous, paroicous or rarely autoicous (not in Victoria), synoicous or polyoicous. Asexual reproduction by rhizoidal tubers (not in Victoria) or more frequently by solitary or clustered gemmae in leaf axils near stem apices (not in Victoria). Scattered plants among other moss or turves on soil, rock or rarely logs. Stem erect, simple or forked at base or sparingly and irregularly branched by innovation when fertile, covered, sometimes sparsely, with rhizoids; central strand present. Leaves elliptic or ovate to lanceolate or linear-lanceolate (not in Victoria), erect and imbricate to erect-spreading when moist, scarcely altered when dry, decurrent or not; apex acuminate or acute; costa terminating in the apical half well below apex, subpercurrent, percurrent or rarely shortly excurrent; margin entire to serrate near apex, entire to serrulate near base, plane or recurved, without a border; laminal cells oblong-hexagonal or rhomboid to linear, sometimes longer and narrower toward margin, usually becoming shorter and quadrate to rectangular at base, smooth, unistratose; alar cells not differentiated. Perichaetia terminal. Perichaetial leaves enlarged compared to vegetative leaves. Capsule erect to pendent, curved or straight, clavate, pyriform, ovoid to cylindric or sometimes urceolate, with a short or long apophysis, without an annulus or with a revoluble annulus. Calyptra cucullate, smooth, glabrous. Operculum convex, conic or apiculate to short-rostrate. Peristome double or rarely absent (not in Victoria); endostome well-developed and around half length to same height as exostome, or rarely rudimentary (not in Victoria) or absent (not in Victoria), with a low to high basal membrane; cilia present and well-developed, or rudimentary or absent.
Worldwide with around 127 species, but mostly in montane habitats and most diverse in northern temperate regions (Shaw 2006); five species in Victoria.
Pohlia is resolved into groups of species that are possibly closer related to other genera than they are to other Pohlia groups in phylogenies based on chloroplast and nuclear DNA sequences, suggesting that the circumscription of Pohlia requires revision (e.g. Guerra et al. 2011; Niu et al. 2018; Ohgue et al. 2021). While the extent of species sampling for these phylogenies is so far low, some morphological basis to the grouping of species in molecular phylogenies is emerging that provides promise for future revision. For instance, one group comprises two species, including the Victorian P. wahlenbergii (F.Weber & D.Mohr) A.L.Andrews, that belong to the section Apalodictyon and differs from other Pohlia by having capsules without annuli and with immersed stomata (Guerra et al. 2011; Niu et al. 2018; Ohgue et al. 2021). All the species that produce axillary bulbils that have been included in molecular phylogenies also form a group (Guerra et al. 2011; Niu et al. 2018; Ohgue et al. 2021).
 Spinning
SpinningGuerra, J.; Jiménez-Martínez, J.F.; Cano, M.J.; Jiménez-Fernández, J.A. (2011). A contribution to the phylogenetic study of Mielichhoferiaceae-Mniaceae (Bryophyta) based on molecular sequence data. Nova Hedwigia 93: 47–56.
Niu, J.-Y.; Li, L.; Shi, S.; Li, H.; Wang, X.-R.; Liu, Y.-Y.; Niu, Y.-L.; Zhao, J.-C. (2018). Phylogenetic analysis of the genus Pohlia (Bryophyta, Bryaceae) using chloroplast and nuclear ribosomal DNA. *Phytotaxa * 351: 141–153.
Ohgue, T.; Akiyama, H.; Suzuki-Azuma, H.; Nagamasu, H. (2021). Phylogenetic study of the genus Pohlia (Mielichhoferiaceae, Bryophyta) based on chloroplast DNA sequences. Bryophyte Diversity and Evolution 44: 48–60.
Shaw, A.J. (2006). A revision of the moss genus Pohlia Hedw. (Mniaceae) in Australia. Systematic Botany 31: 247–257.