Rhabdoweisiaceae
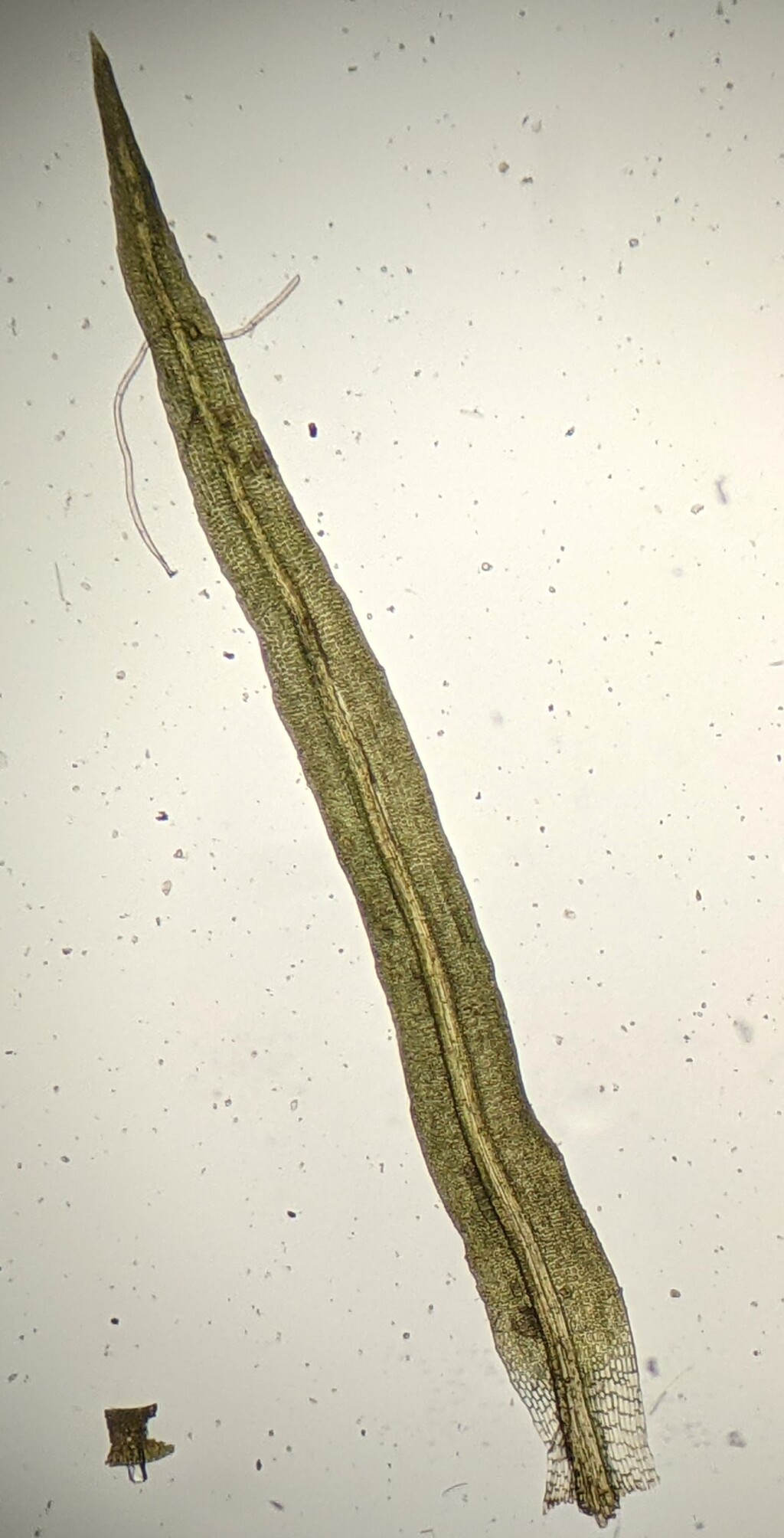
Autoicous, rarely dioicous. Asexual reproduction occasionally by abaxial leaf (not in Victoria) or rhizoidal gemmae (not in Victoria). Tufts, turves, cushions (not in Victoria) or mats on rocks, soil (not in Victoria) or epiphytic (not in Victoria), sometimes aquatic. Stem erect, simple or forked, sometimes moderately branched, glabrous to tomentose (not in Victoria) at base; central strand present, rarely absent (not in Victoria). Leaves arranged around stem so facing all directions, monomorphic, squarrose (not in Victoria) to erect-spreading, falcate-secund or rarely contorted (not in Victoria) when moist, little altered to becoming erect (not in Victoria), contorted (not in Victoria) or crisped (not in Victoria) when dry, with (not in Victoria) or without a sheathing base; apex acuminate or acute (not in Victoria), sometimes with a hairpoint; costa subpercurrent (not in Victoria), percurrent or excurrent, rarely weakly differentiated in parts (not in Victoria) or absent (not in Victoria); margin entire to serrate, recurved (not in Victoria), plane or slightly incurved toward apex, sometimes with a border near base (not in Victoria); laminal cells quadrate, short-rectangular, rarely linear, smooth or mamillose (not in Victoria), unistratose or partly bi- (not in Victoria) or pluristratose (not in Victoria); alar cells undifferentiated or differentiated, quadrate to short-rectangular, brownish or not coloured, sometimes inflated, unistratose or bistratose. Acrocarpous. Capsules exserted, rarely emergent (not in Victoria) or immersed (not in Victoria), erect to horizontal, rarely pendent (not in Victoria), straight or curved, operculate. Calyptra cucullate, rarely mitrate (not in Victoria), smooth, glabrous. Operculum conic (not in Victoria) or rostrate. Peristome a single series of 16 teeth, bifid in upper half or split to near base, rarely entire (not in Victoria) or absent (not in Victoria).
15 genera and 85 species (Fedosov et al. 2021); three genera and species in Victoria.
The Rhabdoweisiaceae in the circumscription followed here is a heterogenous assemblage of taxa, that while monophyletic is not easily defined morphologically and is hard to distinguish from related families of the Dicranales. This is epitomised by several of its species belonging to genera that have now been split into two genera because in their previous circumscriptions, these genera had species that were placed by phylogenetic analyses of chloroplast and mitochondrial DNA in separate families (Hedderson et al. 2004; Werner et al. 2013; Fedosov et al. 2017, 2021). These include Pseudoblindia, that is largely composed of former Blindia (Seligeriaceae) species, Eucamptodon, that previously included the Victorian E. muelleri Hampe & Müll.Hal., now placed in Dicranaceae as Hollia myrmecoa Sieber ex Hoppe, and Dicranoweisia, that previously included the Victorian species Dicranowiesia antarctica (Müll.Hal.) Kindb., now included in Hymenoloma (Hymenolomataceae). Convergent evolution of morphology associated with adaptation to epiphytic or aquatic environments in Rhabdoweisiaceae and other lineages with a single peristome is partly responsible for the lack of morphological difference from other families (Fedosov et al. 2021). When taxa of these specialised habitats are ignored, the remaining taxa of Rhabdoweisiaceae are generally distinguished by a combination of leaves that become curved to crisped or contorted when dry, subquadrate laminal cells in the apical half of the leaf, costae with developed stereid bands and a peristome or 16 split teeth with vertical striae on their outer surface (Fedosov et al. 2021).
Several Rhabdoweisiaceae genera have previously been included in the Dicranaceae, including the Victorian Kiaeria. However, these genera and other taxa included here in Rhabdoweisiaceae, form a lineage that is closer related to other well-established families than to Dicranaceae (Hedderson et al. 2004; La Farge et al. 2000, 2002; Stech et al. 2012; Fedosov et al. 2021).
Amphidium, which was traditionally included in Rhabdoweisiaceae, has been shown to be less closely related to Rhabdoweisiaceae than some other well-established families (La Farge et al. 2000; Stech et al. 2012; Fedosov et al. 2021). Consequently, Amphidium has been segregated as their own family (Stech & Frey 2008).
 Spinning
SpinningFedosov, VE; Fedorova, A.V.; Ignatova, E.A.; Ignatov, M.S. (2017). A revision of the genus Seligeria (Seligeriaceae, Bryophyta) in Russia inferred from molecular data. Phytotaxa 323: 27–50.
Fedosov, V.E.; Fedorova, A.V.; Larraín, J.; Santos, M.B.; Stech, M.; Kučera, J.; Brinda, J.C.; Tubanova, D.Y.; Von Konrat, M.; Ignatova, E.A.; Ignatov, M.S. (2021). Unity in diversity: phylogenetics and taxonomy of Rhabdoweisiaceae (Dicranales, Bryophyta). Botanical Journal of the Linnean Society 195: 545–567.
Hedderson, T.A.; Murray, D.J.; Cox, C.J.; Nowell, T.L. (2004). Phylogenetic relationships of haplolepideous mosses (Dicranidae) inferred from rps4 gene sequences. Systematic Botany 29: 29–41.
La Farge, C.; Mishler, B.D.; Wheeler, J.A.; Wall, D.P.; Johannes, K. (2000). Phylogenetic relationships within the haplolepidous mosses. The Bryologist 103(2): 257–276.
La Farge, C.; Shaw, A.J.; Vitt, D.H. (2002). The circumscription of the Dicranaceae (Bryopsida) based on the chloroplast regions trnL-trnF and rps4. Systematic Botany 27: 435–452.
Stech, M.; Frey, W. (2008). A morpho-molecular classification of the mosses (Bryophyta). Nova Hedwigia 86: 1–21.
Stech, M.; McDaniel, S.F.; Hernández-Maqueda, R.; Ros, R.M.; Werner, O.; Muñoz, J.; Quandt, D. (2012). Phylogeny of haplolepideous mosses – challenges and perspectives. Journal of Bryology 34: 173–186.
Werner, O.; Rams, S.; Kučera, J.; Larraín, J.; Afonina, O.M.; Pisa, S.; Ros, R.M. (2013). New data on the moss genus Hymenoloma (Bryophyta), with special reference to H. mulahaceni. Cryptogamie, Bryologie 34(1): 13–30.